
Взаимодействие человека и животных с окружающей средой может осуществляться только через движение. Существует два вида двигательных функций: поддержание положения тела (позы) и собственно движения. Среди собственно движений следует различать элементарные движения и действия.
Последние – действия, представляют собой совокупность элементарных движений, объединенных единой целью в некоторую функциональную систему. Каждое конкретное действие включено в состав поведенческого акта. Оно является шагом к удовлетворению той потребности, которая вызвала данное поведение, и решает свою промежуточную задачу.
Произвольные движения человека – это сознательно регулируемые движения, которые осуществляются в отличие от непроизвольных с определенной целью.
Произвольные двигательные акты связаны с функцией пирамидального и экстрапирамидального трактов, связывающих кору головного мозга и подкорковые структуры со спинальным моторным механизмом (мотонейронами спинного мозга).
Впервые наличие прямых эфферентных и обратных афферентных связей между нервными структурами спинного мозга и мышечным аппаратом была показана в работе Чарльза Белла, который описал нервный круг: один нерв несет импульсы мозга к мышце, другой приносит ощущения состояния мышцы к мозгу.
Морфо- и функциогенез мозга || лекция 7-8/10 || Наталья Чурило
Позже И.М. Сеченов выделил понятие «темное мышечное чувство», которое по его убеждению порождает представление о скорости движения и отношениях предметов в пространстве. Рецепторный аппарат мышц, сухожилий и суставов, образующих «мышечное чувство» Ч.С. Шеррингтон назвал проприорецепторами.
Шеррингтон считал, что главная функция проприорецепторов – давать информацию о собственных движениях организма. В настоящее время выделено два типа проприорецепторов: рецепторы растяжения и рецепторов натяжения мышечных волокон. В эксперименте в 1909 году И.П. Павлов установил, что моторная кора получает сенсорные сигналы от рецепторов мышц и суставов и ввел понятие двигательного анализатора. В результате чего моторная кора стала рассматриваться как центральный аппарат построения движений.
Обобщив перечисленные факты, П.К. Анохин применил понятие обратная связь или обратная афферентация. Таким образом, основной принцип в механизме координации двигательных реакций является циклическая нервная связь и кольцевая структура управления или рефлекторное кольцо по Н.А. Бернштейну.
Однако не все двигательные акты осуществляются по кольцевому механизму – движения, базирующиеся на врожденных координациях, в меньшей степени требуют обратной связи, в то время как приобретаемые движения всецело зависят от соматической афферентации с двигательного аппарата. Движения, осуществляющиеся без помощи проприоцептивной обратной связи, выполняются по жесткой программе. Но даже эти движения требуют сличения с определенным сенсорным образом. П.К. Анохин для обозначения механизма сличения выделил в функциональной системе двигательного акта специальный блок сличения, который назвал акцептор результата действия.
Центры коры полушарий головного мозга. Организация коры.
В управлении движением различают стратегию и тактику. Основу стратегии движения определяет конкретная мотивация (биологическая, социальная и др.). Именно на ее основе определяется цель поведения, т. е. то, что должно быть достигнуто. В структуре поведенческого акта цель закодирована в акцепторе результатов действия.
В отношении движения это выглядит как формирование двигательной задачи, т. е. того, что следует делать. Под тактикой понимают конкретный план движений, т. е. то, как будет достигнута цель поведения, с помощью каких двигательных ресурсов, способов действия.
Таким образом, можно выделить два механизма управления движением:
— посредством центральных моторных программ;
— с помощью обратной афферентации.
Впервые представление о центральных моторных программах было дано в работе Ч. Шеррингтона, который занимался изучением функций мотонейронов спинного мозга собак при осуществлении двигательных актов ходьбы и чесания. Производя перерезки участков спинного мозга, он показал, что механизмы, ответственные за некоторые двигательные акты локализованы в спинном мозге. Через несколько месяцев после перерезки у собак удавалось вызвать чесательный рефлекс на механическое раздражение кожи. На основании чего была заложена концепция о «запускаемом движении» с помощью центральных программ. Позже было показано, что ходьба животного – перемещение в окружающей среде при помощи координированных движений конечностей – это также запрограммирована на уровне спинного мозга.
Программы центральных моторных программ широко представлены в различных структурах мозга и ЦНС хранит как врожденные, так и приобретенные программы выработанных действий. Например, дыхательные, глотательные движения управляются врожденными программами, а профессиональные, спортивные и иные навыки осуществляются по программам, формирующимся в течение жизни индивида. При определенном опыте человека эти программы выполняются автоматически, а обратная афферентация перестает играть существенную роль. Обратная афферентация становится нужна при переделке – корригировании (корректировке) навыка.
У позвоночных центральные программы запускаются управляющими центрами, а у беспозвоночных, как правило, командными нейронами, впервые выделенными в 1968 г. К. Вирсма. Однако сигнал от командного нейрона или управляющего центра играет лишь пусковую роль. Он не несет информации о том, как построить соответствующее движение. Структура движения заложена во взаимосвязях мотонейронов, с которыми он связан.
Рассмотрим несколько примеров поведения у низших животных:
— рефлекс бегства у рака, легко вызываемый тактильным, вибрационным или звуковым раздражением, реализуется через возбуждение одного гигантского командного нейрона, связанного с тремя двигательными мотонейронами;
— в ответ на стимуляцию рыба уплывает, совершая быстрые и повторяющиеся движения хвостом и головой; этот двигательный акт запускается парой командных нейронов – маутнеровских клеток, расположенных в продолговатом мозгу;
— у таракана есть командный нейрон, запускающий бег насекомого, если его лапы касаются опоры, если же лапы находятся в воздухе запускается команда полета.
Существует два типа командных нейронов: одни лишь запускают двигательную программу, не участвуя в ее дальнейшем осуществлении, другие поддерживают или видоизменяют двигательную программу. Первый тип нейронов получил название триггеры. Второй тип нейронов получил название воротных нейронов.
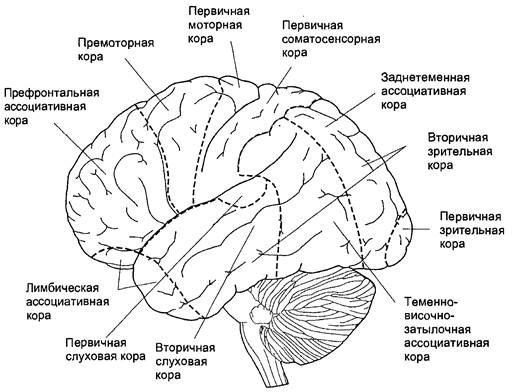
Рисунок 3 – Схема двигательной коры большого мозга
Ведущая роль в построении новых моторных программ у высших животных и человека принадлежит передним отделам коры больших полушарий – премоторной и префронтальной коре. Роль передних отделов коры больших полушарий показана в работах А.С. Батуева, проводившего эксперименты с использованием микроэлектородов, регистрирующих активность отдельных нейронов у обезьян.
В экспериментах были обнаружены три группы нейронов, последовательно вовлекаемых в процесс осуществления условнорефлекторного двигательного навыка. Все три популяции клеток были обнаружены в теменной и особенно в лобной коре.
Первая группа клеток реагирует на условные сигналы, поэтому их отнесли к сенсорным нейронам. Вторая группа клеток реагирует на время действия сигнала, а третья группа в пусковой период. Считают, что сенсорные нейроны первой группы передают информацию второй группе, которые принадлежат кратковременной памяти. Третья же группа – это нейроны моторных программ, они получают информацию от нейронов памяти и запускают хорошо отработанную двигательную реакцию.
Многие исследователи считают, что хранение части подобных моторных программ осуществляется структурами мозжечка. Кроме того, в процессе индивидуальной жизни мозжечок обучается различным программам движения и сохраняет их. В этом процессе важную роль играют клетки Пуркинье.
Мозжечок получает информацию о планируемом корой большого мозга движении, вносит коррекцию в план будущего движения и посылает ее в кору большого мозга. Быстрые мелкие точные движения осуществляются без сличения корой с импульсами от проприорецепторов. Мозжечок обучается также корригировать (изменять) выполняемое движение, играет важную роль в регуляции тонуса мышц, поддержании позы, координации движения.
В качестве другой центральной структуры, связанной с хранением центральных моторных программ рассматриваются базальные ганглии. Базальные ганглии имеют отношение к хранению программ врожденных двигательных актов и быстрых стереотипных движений. Врожденные программы имеются также в спинном мозге (шагательный рефлекс) и в стволе мозга (дыхания, глотания, выпрямительный рефлекс – восстановление нарушенной позы, ориентировочный рефлекс).
Инициация движения происходит следующим образом. В моторной коре строится конечный и конкретный вариант моторного управления движением. Моторная кора использует оба принципа управления: контроль через систему обратной афферетации и через механизм центрального программирования. Это достигается тем, что к ней сходятся сигналы от мышечной активности, от сенсомоторной, зрительной и других отделов коры, которые и используются для моторного контроля коррекции движения. Кроме того, к моторной коре приходят сигналы, связанные с программированием движения из передних отделов коры и подкорки.
Моторная кора получает также информацию из структур, имеющих отношение к хранению двигательных программ. Так, в опытах с регистрацией нейронной активности у обезьян было показано, что при выполнении ими заученного движения активность нейронов зубчатого ядра мозжечка на 10 мс опережает изменение активности нейрона в моторной коре, которое предшествует появлению мышечного движения.

Рисунок 2 — Схема инициации двигательного акта (по Г. Могенсону, 1977, с изменениями)
В моторную кору поступают также сигналы от базальных ганглиев – структура, которая ответственна за хранение главным образом двигательных программ врожденного поведения (пищевого, питьевого и др.), стереотипных движений. Клетки базальных ганглиев, так же как и мозжечка, разряжаются до начала движений, совершаемых животными в ответ на сигнал за 50 – 150 мс. В хвостатом ядре были найдены нейроны, у которых возбуждение опережает условную двигательную пищевую и оборонительную реакции. Эти реакции у нейронов появляются с выработкой условного рефлекса и исчезают с его утешением. Сигналы от базальных ганглиев и мозжечка распространяются не только вверх к коре, но и вниз, по нисходящему пути к спинному мозгу (так называемый экстрапирамидный путь).
Согласно работам шведского нейрофизиолога Г. Могенсона, все процессы управления движением включают три блока (рассмотрим рисунок 2) и соответствующие им три фазы:
1) блок инициации движения, включающий лимбическую систему с прилегающим ядром (n. accumbens), и ассоциативную кору;
2) блок программирования движения, включающий мозжечок, базальные ганглии, моторную кору, таламус (как посредник между ними), а также спинальные и стволовые генераторы;
3) исполнительный блок, охватывающий мотонейроны и двигательные единицы.
Таким образом, произвольные движения, так же как и непроизвольные, являются результатом сопряженной работы лимбической системы и ассоциативной коры. Двигательная система работает как многоуровневая. На каждом уровне имеется своя «ведущая афферентация» и собственный тип регулируемых движений.
Однако произвольные движения без пирамидной системы осуществляться не могут, а непроизвольные – могут. Непроизвольные движения запускаются посредством возбуждения экстеро- и проприорецепторов, могут осуществляться с помощью любого уровня ЦНС, в том числе и на уровне спинного мозга. Произвольные движения инициируются ассоциативной корой и лимбической системой с помощью механизмов памяти.
Источник: studopedia.su
43 Механизмы управления движением
Строится на двух основных принципах — принципе сенсорных коррекций текущего движения по цепи обратной связи и принципе прямого программного управления. Последний особенно важен для тех случаев, когда имеются быстрые изменения в системе и это ограничивает возможность сенсорных коррекций.
Существование двух механизмов управления движением: посредством центральных моторных программ и с помощью обратной афферентации, которая используется для непрерывного контроля и коррекции выполняемого движения. Для многих видов движения управление может осуществляться одновременно двумя механизмами при разном их соотношении для движений, различающихся сложностью и уровнем организации.
Сенсорные раздражения не только могут запускать движение, но и выполнять корректирующую функцию. Обратная афферентация, сигнализирующая о результатах действия, сопоставляется с программой движений и служит уточнению координат цели и траектории движения.
«проприоцепция» — обозначение сенсорных входов, которые возбуждаются во время движения. Главная функция проприоцепторов — давать информацию о собственных движениях организма. Выделено два типа проприоцепторов: рецепторов растяжения, активирующихся при растяжении мышц, и рецепторов напряжения, которые чувствительны к силе сокращения мышцы.
Моторная кора получает сенсорные проекции от рецепторов мышечной и суставной чувствительности. Понятие двигательного анализатора, выполняющего функцию восприятия сигналов от тела.
Изменения в мышце, возникающие при движении, возбуждают чувствительные окончания проприоцепторов, а эти проприоцептивные сигналы, достигая моторных центров, вносят изменения в эффекторный поток, т. е. в физиологическое состояние мышцы. Таким образом, перед нами «. не рефлекторная дуга, а другая форма взаимоотношений между афферентным эффекторным процессом, характерная для всех координационных процессов — рефлекторное кольцо». Рефлекторное кольцо представляет собой фундаментальную форму протекания двигательного нервного процесса.
Первые представления о том, что движения могут управляться не только по типу непрерывной коррекции, но и через механизм центральных моторных программ, также возникли на основе работ Ч. Шеррингтона. Сигналы, идущие от многих различных областей головного мозга, часто сходятся к одним и тем же мотонейронам спинного мозга. «общий конечный путь», который связывает головной мозг с мышечным актом. Механизмы, ответственные за некоторые двигательные акты, локализованы в спинном мозге.
Программы цепных двигательных актов широко представлены в различных структурах мозга. Центральная нервная система хранит центральные программы как врожденных, так и выработанных действий. У позвоночных центральные программы запускаются управляющими центрами.
Ведущая роль в построении новых моторных программ принадлежит передним отделам коры больших полушарий (премоторной, префронтальной коре). Происходит выбор отдельных фрагментов из имеющихся программ как врожденных форм поведения, так и приобретенных двигательных автоматизмов для их последующей интеграции в новую моторную программу.
Мозжечок обучается различным программам движения, а затем и сохраняет их. В нем хранятся программы сложных и автоматически выполняемых двигательных актов, которые сформировались при жизни животного и человека. В качестве другой центральной структуры, связанной с хранением центральных моторных программ, исследователи рассматривают базальные ганглии — структуры, расположенные в глубине белого вещества полушарий.
Источник: studfile.net
Роль коры больших полушарий
В РЕГУЛЯЦИИ ТОНУСА И УПРАВЛЕНИИ ДВИЖЕНИЯМИ.
«Третий этаж» или уровень регуляции движений — это кора больших полушарий, которая организует формирование программ движений и их реализацию в действие. Зарождающийся в ассоциативных зонах коры замысел будущего движения поступает в моторную кору. Нейроны моторной коры организуют целенаправленное движение с участием БГ, мозжечка, красного ядра, вестибулярного ядра Дейтерса, ретикулярной формации, а также – с участием пирамидной системы, непосредственно воздействующей на альфа-мотонейроны спинного мозга.
Корковое управление движениями возможно лишь при одновременном участии всех моторных уровней. Двигательная команда, передаваемая из коры головного мозга, оказывает воздействие через более низкие моторные уровни, причём каждый из них вносит свой вклад в окончательную двигательную реакцию. Без нормальной деятельности нижележащих моторных центров корковое моторное управление было бы несовершенным.
Сейчас уже многое известно о функциях моторной коры. Её рассматривают как центральную структуру, управляющую самыми тонкими и точными произвольными движениями. Именно в моторной коре строится конечный и конкретный вариант моторного управления движениями. Моторная кора использует два принципа управления движениями: контроль через петли обратной сенсорной связи и через механизмы программирования. Это достигается тем, что к ней сходятся сигналы от мышечной системы, от сенсомоторной, зрительной и других отделов коры, которые и используются для моторного контроля и коррекции движения.
Афферентные импульсы к моторным зонам коры поступают через моторные ядра таламуса. Через них кора связана с ассоциативными и сенсорными зонами самой коры, с подкорковыми базальными ганглиями и мозжечком.
Моторная область коры регулирует движения с помощью эфферентных связей трёх типов: а) прямо на мотонейроны спинного мозга через пирамидный тракт, б) косвенно при помощи связи с нижележащими двигательными центрами, в) ещё более косвенная регуляция движений осуществляется путём влияния на передачу и обработку информации в чувствительных ядрах мозгового ствола и таламуса.
Как уже говорилась, сложную моторную деятельность, тонкие координированные действия определяют моторные области коры, от которых к стволу и спинному мозгу направляются два важных пути: кортикоспинальный и кортикобульбарный, которые иногда объединяют под названием пирамидный тракт. Кортикоспинальный путь, обеспечивающий управление мышцами туловища и конечностей, заканчивается либо прямо на мотонейронах, либо на интеронейронах спинного мозга. Кортикобульбарный путь осуществляет контроль двигательных ядер черепно-мозговых нервов, управляющих мышцами лица и движениями глаз.
Пирамидный тракт является самым большим нисходящим моторным путём; он образован приблизительно одним миллионом аксонов, больше половины которых принадлежит нейронам, которые называются клетки Беца или гигантские пирамидные клетки. Они расположены в V слое первичной моторной коры в области прецентральной извилины.
Именно от них берёт начало кортикоспинальный путь или так называемая пирамидная система. Через посредство вставочных нейронов или путём прямого контакта волокна пирамидного тракта образуют возбуждающие синапсы на мотонейронах сгибателей и тормозные –на мотонейронах разгибателей в соответствующих сегментах спинного мозга. Спускаясь к мотонейронам спинного мозга, волокна пирамидного тракта отдают многочисленные коллатерали к другим центрам: красному ядру, ядрам моста, ретикулярной формации ствола мозга, а также к таламусу. Эти структуры связаны с мозжечком. Благодаря связям моторной коры с двигательными подкорковыми центрами и мозжечком, она участвует в обеспечении точности выполнения всех целенаправленных движений — как произвольных, так и непроизвольных.
Пирамидный путь частично перекрещивается, поэтому инсульт или иное повреждение правой моторной зоны вызывает паралич левой половины тела, и наоборот
До сих пор можно встретить, наряду термином пирамидная система, ещё один: экстрапирамидный путь или экстрапирамидная система. Этот термин применялся для обозначения других двигательных путей, идущих от коры к двигательным центрам. В современной физиологической литературе термин экстрапирамидный путь и экстрапирамидная система не используется.
Нейроны в двигательной коре, также как и в сенсорных областях, организованы в вертикальные колонки Корковая моторная (другое название – двигательная) колонка – это небольшой ансамбль двигательных нейронов, которые контролируют группу связанных между собой мышц. Как сейчас полагают, их важная функция состоит не просто в том, чтобы активировать те или другие мышцы, а в том, чтобы обеспечивать определенное положение сустава. В несколько общей форме можно сказать, что кора кодирует наши движения не путем приказов о сокращении отдельных мышц, а путем команд, обеспечивающих определенное положение суставов. Одна и та же группа мышц может быть представлена в разных колонках и может участвовать в разных движениях
Пирамидная система является основой наиболее сложной формы двигательной активности — произвольных, целенаправленных движений. Кора больших полушарий является субстратом для обучения новым видам движений (например, спортивным, производственным и т.д.). В коре хранятся сформированные в течение жизни программы движений,
Ведущая роль в построении новых моторных программ принадлежит передним отделам КБП (премоторной, префронтальной коре). Схема взаимодействия ассоциативных, сенсорных и моторных областей коры при планировании и организации движений представлена на рисунке 14.
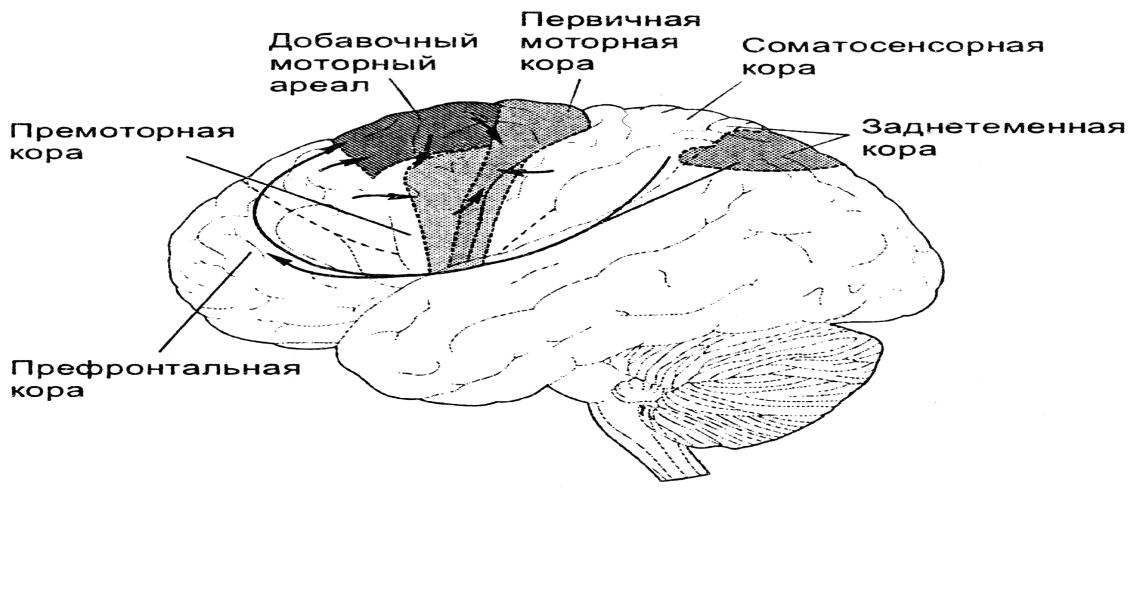
Рисунок 14. Схема взаимодействия ассоциативных, сенсорных и моторных областей при планировании и организации движений
Планировать предстоящие действия начинает префронтальная ассоциативная кора лобных долей на основе информации, поступающей, в первую очередь, от заднетеменных областей, с которыми её связывает множество нейронных путей. Выходная активность префронтальной ассоциативной коры адресована премоторным или вторичным моторным областям, которые создают конкретный план предстоящих действий и непосредственно готовят моторные системы к движению.
К вторичным двигательным областям относятся премоторная кора и добавочный моторный ареал (добавочная моторная область). Выходная активность вторичной моторной коры направлена к первичной моторной коре и к подкорковым структурам. Премоторная область контролирует мышцы туловища и проксимальные отделы конечностей. Эти мышцы особенно важны в начальной фазе выпрямления тела или движения руки к намеченной цели. В отличие от этого, добавочный моторный ареал участвует в создании модели двигательной программы, а также программирует последовательность движений, которые выполняются билатерально (например, когда надо действовать обеими конечностями).
Вторичная моторная кора занимает в иерархии двигательных центров главенствующее над первичной моторной корой положение: во вторичной коре движения планируются, а первичная — этот план выполняет.
Первичная моторная кора обеспечивает простые движения. Она расположена в передних центральных извилинах мозга. Исследования на обезьянах показали, что в передней центральной извилине имеются неравномерно распределенные зоны, управляющие различными мышцами тела. В этих зонах мышцы тела представлены соматотопически, то есть каждой мышце соответствует свой участок области (двигательный гомункулус) (рис 15).
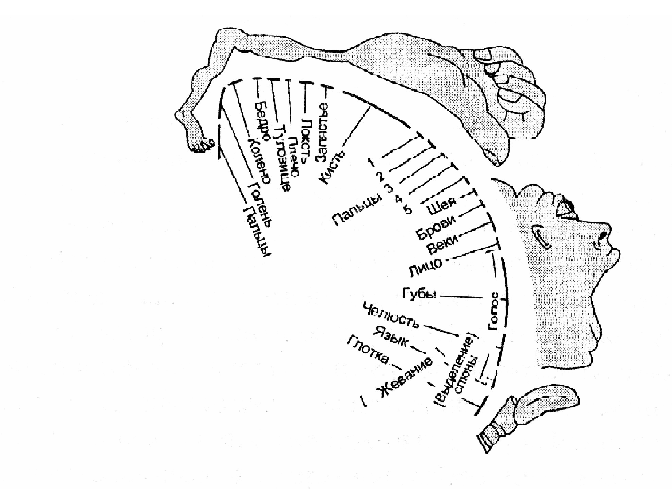
Рисунок 15. Соматотопическая организация первичной моторной коры — двигательный гомункулус
Как показано на рисунке, самое большее место занимает представительство мышц лица, языка, кистей рук, пальцев — то есть тех частей тела, которые несут наибольшую функциональную нагрузку и могут совершать самые сложные, тонкие и точные движения, и в то же время сравнительно мало представлены мышцы туловища и ног.
Моторная кора управляет движением, используя информацию, поступающую как по сенсорным путям от других отделов коры, так и от генерируемых в ЦНС моторных программ, которые актуализируются в базальных ганглиях и мозжечке и доходят до моторной коры через таламус и префронтальную кору. Полагают, что в БГ и в мозжечке уже заложен механизм, который может актуализировать хранящиеся в них двигательные программы. Однако, для приведения в действие всего механизма необходимо, чтобы в эти структуры поступил сигнал, который послужил бы начальным толчком процесса. По-видимому, существует общий биохимический механизм актуализации моторных программ в результате роста активности дофаминергических и норадренергических систем в мозге.
Согласно гипотезе, высказанной П. Робертсом, актуализация моторных программ происходит вследствие активации командных нейронов. Существует два типа командных нейронов. Одни из них лишь запускают ту или иную двигательную программу, но не участвуют в её дальнейшем осуществлении. Эти нейроны называются нейроны-триггеры.
Командные нейроны другого типа получили название воротных нейронов. Они поддерживают или видоизменяют двигательные программы лишь будучи в состоянии постоянного возбуждения. Такие нейроны обычно управляют позными или ритмическими движениями. Сами командные нейроны могут контролироваться и затормаживаться сверху. Снятие торможения с командных нейронов повышает их возбудимость и тем самым высвобождает «предпрограммированные» цепи для той деятельности, для которой они предназначены
В заключение следует отметить, что двигательные (моторные) области коры больших полушарий служат последним звеном, в котором образованный в ассоциативных и других зонах (а не только в моторной зоне) замысел превращается в программу движения. Главной задачей моторной коры является выбор группы мышц, ответственных за выполнение движений в каком-либо суставе, а не непосредственная регуляция силы и скорости их сокращения. Эту задачу выполняют нижележащие центры вплоть до мотонейронов спинного мозга. В процессе выработки и реализации программы движения моторная область коры получает информацию от БГ и мозжечка, которые посылают к ней корректирующие сигналы.
Запомните:
КОРА БОЛЬШИХ ПОЛУШАРИЙ:
| —регулирует силу спинномозговых и стволовых двигательных рефлексов; |
| —участвует в формировании и хранении программ сложных врожденных и всех приобретенных движений; |
| —обеспечивает выполнение целенаправленных произвольных двигательных актов. |
| —в коре формируется замысел или цель движения, происходит выбор программ движения, а также запуск сложных видов движений; |
Заметим, что пирамидные, руброспинальные и ретикулоспинальные пути активируют преимущественно флексорные, а вестибулоспинальные — преимущественно экстензорные мотонейроны спинного мозга. Дело в том, что флексорные двигательные реакции являются основными рабочими двигательными реакциями организма и требуют более тонкой и точной активации и координации. Поэтому в процессе эволюции большинство нисходящих путей специализировалось по активации именно флексорных мотонейронов.
Источник: studopedia.org